












酵素やホルモン・サイトカインなどの活性を測定する時の注意点
−濃度と時間の大切さ−
堀池 喜八郎 生化学第一講座
■生物活性(あるいは応答)の表し方
●酵素活性の場合=初速度
酵素の働きの強さ(触媒能),すなわち,その触媒する反応の速度(基質の減少速度-d[S]/dtや生成物の増加速度d[P]/dt)をその酵素の活性(enzyme activity)という.酵素活性を測定することはその酵素の触媒能の程度や酵素の有効量を知るために必須である.
酵素反応の速度は,反応時間が経過するにつれて減少する(平衡に近づく)ので,活性測定のためには反応の初期での速度(反応時間を0に外挿した時点での反応速度,初速度initial velocity)が用いられる.初速度を反応速度の指標とすることの意義は,生成物による阻害や反応中に起こりうる酵素の失活の効果を除いて,取り扱いを単純化することである.
ここで次のような疑問をもつかもしれない.「[P] = 0という点での初速度υを考えている.生成物ができてもいないのに反応速度は測れるのか?」と.答は「できる」.y = 2 xという直線を考える.yは生成物の濃度,xは時間であると思いながら,この直線の傾きを考えよう. x = 0でのyの値は0であるが,x = 0での傾きは2である.言い換えれば,微分dy/dxのx = 0での値は2である.x = 0だからといって,微分量も0になっているわけではない.反応速度は微分量d[P]/dtである.[P] = 0でもυ= d[P]/dtは0ではない.(実際には、[P]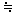 0である)
0である)
●ホルモンやサイトカインなどの生理活性物質の場合=平均の応答速度
生理活性物質の作用を調べる場合も,それら化合物の添加後,適当な時間がたってから(ある時間incubationしてから),目的に応じて特定の化合物を分析・定量している.多くの酵素反応のように連続的に変化を追跡することはまれである.
当然のことながら,経時変化の十分な検討はすでになされているという前提のもとに,ある任意の勝手に決めた一定時間後に分析している.測定までの時間経過中に,応答にlagはあったのか,そうではなく直線的に変化したのか,あるいはピーク(極大値)を示したのか.もし経時変化の全体的パターンが不明なら,結論を誤るかもしれない.なぜなら,一定時間当たりの変化量(平均の応答速度)だけを観測しているだけだから.このことをしっかりと肝に銘じておこう.
●絶対測定と相対測定
例:ある臓器で物質Xを定量し(絶対測定),それをコントロールに対する百分率で表し(相対測定),処置の効果を判定した.ところが,コントロールでの物質Xの絶対的含有量(〜nmol/g wet weight)に疑義があった.
■試薬の混合順序が活性に影響することがある
●難溶性の試薬
水に難溶性の試薬(ブロッカーなど)を溶かすため,detergentや有機溶媒(エタノール,アセトンなど)を用いることがある.
このdetergentや有機溶媒がタンパク質と相互作用し,タンパク質の立体構造が変化し,その結果,活性も変わることがある.あるいは,反応容器に難溶性試薬が強く吸着し有効濃度が減少したり,次の実験のときにそれが吸着面から時間とともにゆっくりと溶け出すことがある.
●試薬間での反応や相互作用
試薬同士が反応する.塩の形成など(Caイオンとリン酸イオン)
■定数を求めるときの濃度の選び方
●比活性あるいはstandard assayの実験条件の決め方
酵素を取り扱う際,しばしば比活性を測定する必要にせまられる.実験には必ず誤差が伴うが,これをできる限り少なくしなければならない.そのために以下のことに留意する.
●
実験者には(無意識下で)線形プロットをしているという意識があり,手には直定規をもっている.その物差しをデータポイントにあてる.この時点であなたは,この酵素反応はMichaelis-Menten型(allostericな挙動はない)としている.なぜなら直線をひいているのだから.特に1/υ−1/[S]plotの場合,直線に読みとってしまう.このplotは危険なplotである.統計的にも各基質濃度での誤差が一様でなく,allostericな振る舞いを見落としてしまう.
Km や Vmax を求めるときは,Eisenthal−Cornish-Bowdenのdirect linear plotやHanes-Woolf plot ([S]/v-[S] plot)を用いる.Lineweaver-Burk plotは阻害のパターンの判定などに用いて,定数の算出には用いない.
例1:L-B plotでplotが横軸(1/S軸)と平行になり,Kmが求められないと,相談を受けた.
例2:L-B plotでplotが上に凸であるのに,直線をあてはめてしまった.
例3:plotを統計解析のソフトで処理(多項式でfitting)し,その理論的根拠を尋ねられた.
■「定数」の意味について
●定数には,実験条件が変わっても不変である定数と,そうでない定数の二種類がある
1)普遍定数(universal constant)
基本法則を表す式において,関与する対象の個々の種類および状態に関係なく,つねに一定の値を保つ定数:アボガドロ定数,ボルツマン定数,プランクの定数,光速度定数(真空中の光速度),…
2)ある一定の条件のもとで一定である定数
比活性,Km,Vmax,速度定数,受容体や酵素へのリガンドの結合定数や解離定数.これらの値は,pH・温度・イオン強度など熱力学的条件が変わると変化する(これら条件の関数である).
●定数や物理量の単位について
M-M式,v=VS/[Km + S],の分母は Km + S であり,Sの単位はモル濃度であるから,Kmの単位も当然モル濃度である.同じ単位でなければ,足すことはできない.分母の単位のモル濃度と分子のSの単位のモル濃度は互いに消去して,M-M式の単位は,vの単位=Vの単位,すなわち,化学反応の速度の単位となる.
阻害剤の解離定数は反応の平衡定数であり,その単位は濃度であるが,平衡定数の単位は濃度とは限らず,濃度の逆数であったり,無名数であったりする.
もう一つの例:気体の状態方程式 PV = nRT の話
■effectorやfactorの添加で活性が上昇したからといって,「活性化」したとは限らない
シグナルが細胞に伝達され,protein kinaseが活性化され,酵素Eがリン酸化されたとする.その結果Km・Vmaxともに低下したとする.この場合,実験で用いる[S]の値によって,活性は上がるか,変わらないか,下がるか,どれでも好きなものすることができる.Eのリン酸化によって,Km とVmax がともに大きくなったときも,全く同じように,[S]の値の選び方によって活性の変化を好きにしうる.
活性化,不活性化はKm とVmax しだいである.したがって,今問題となっている酵素のkinetic parametersの値は測定しておかなければ,結論を誤るかもしれない.
酵素のリン酸化・脱リン酸化に伴って,酵素は活性型や不活性化型に変換する.このようにtextに記載されているが,不活性型酵素で活性が0である,あるいは活性型に比較して,非常に活性が低いという例はほとんどない.活性の差はわずかである.しかし,応答としての代謝の流量は大きく変化する.
■活性化や不活性化を定量的に取り扱って,考えるということ
ある酵素の活性がリン酸化で上がった,あるいは,下がったといって,それだけで代謝の向きは説明できても,その量(代謝の流量)は説明できない.
また,ある反応のアロステリックエフェクターの濃度が増えたといって,流量変化の向きは説明できても,その量的変化はそれだけでは説明できない.
例:解糖系の調節
■扱っているeffectorやfactorと拮抗するfactorを添加してみること
拮抗する作用をもつfactorを量を変えて添加し,実験する.
例:insulinとglucagon
in vivoで,[insulin]=0あるいは[glucagon]=0とはならないので.
■effectorやfactor添加後の時間変化
●ヒステリーシス現象・履歴酵素−もう1つのアロステリック効果(協同現象)
●アロステリック現象の概念の拡張
酵素反応のυ-[S]曲線やタンパク質へのリガンドの結合曲線は,直角双曲線を示すタイプとそれとは異なるタイプ(たとえばS字状)の2つに大別される.非双曲線は,タンパク質の立体構造の変化あるいはその結果としての結合部位間の相互作用に起因し,協同性(cooperativity) あるいはアロステリック効果が認められるという.この現象は生体機能の調節機構の1つとして重要であり,現在ではモノーら(1963年)による原義である“立体構造上(空間的に)異なる部位”をはなれて,広くアロステリー (allostery) またはアロステリズム (allosterism) と呼ばれる.
■生理活性物質(基質・ホルモン・サイトカインなど)の濃度に対する生物応答のパターンは5(+1)種類に分類される
ある物質の濃度(用量)とそれによってひき起こされた反応の強さ(応答)との間の関係は,
これらの曲線は,縦軸に反応の強さをとり横軸に化合物の濃度(遊離濃度)をとって表されるのが普通である.曲線のパターンには次の5つのタイプがある.
■理論式中の生理活性物質の濃度は,ふつう全濃度でなく,遊離の濃度である
●多くの酵素反応の実験では[S][S]t と近似できる
基質の全濃度[S]t は,遊離の濃度[S],酵素に結合しているものの濃度[ES],および生成物の濃度[P]の和で表される.
[S]t = [S] + [ES] + [P]
ふつう酵素活性を測定するときは,酵素の全濃度[E]t に対して基質の全濃度[S]t は十分に大きいので([S]t >>[E]t),たとえすべての酵素に基質が結合したとしても[S]>>[ES]である.さらに反応の初速度を考えているから[P] 0である.
したがって,通常の実験では[S][S]t と近似でき,Michaelis-Mentenの式も次のように書ける.
υ = Vmax[S]/Km+[S] → υ = Vmax[S]t /Km+[S]t
例:基質の全濃度[S]t が,酵素の全濃度[E]t よりも1,000倍以上の桁が高いとしよう.[S]t >>[E]t であり,[S][S]t と近似できる.今,[S]t =1,[E]t =0.001とする.酵素すべてに基質が結合したとすると,遊離の基質濃度[S]は0.999となる.我々の行う通常の実験では有効数字は2桁ないし3桁であるので,[S][S]t と近似できるわけである.
ちなみに,有効数字を3桁にすると,我々は重さ・体積・温度・時間などすべての実験条件を4桁目まで制御しなければならない.普通の生化学の研究室ではかなり困難である.
●多くのホルモンやサイトカインの実験では[L][L]t と近似できない
ホルモン・サイトカインなどの生理活性物質を用いる多くの実験系では,これらリガンドの濃度は酵素反応の場合の基質の濃度に比べ,非常に低い濃度であり,[L][L]t と近似できない場合が多い.注意しよう.
このため,スキャッチャードプロット(Scatchard Plot)などで解析するとき,遊離(非結合)のリガンド濃度を直接的に測定するか,計算で求めなければならない.
●酵素反応の実験で[S][S]t と近似できない場合の計算法
酵素の活性が低く,活性測定の際,大量の酵素を用いる必要があり,実験的に [S]t >>[E]t とできない,つまり[S][S]t とできない場合がある.このときの[S]の求め方を以下に示す.(我々の知っている量は[S]t と,そのときのυの値,それに[E]t だけである)
| [S]t=[S]+[ES]+[P] ∴[S]=[S]t − [ES] |
(初速度 →[P] 0) |
(1) | ||
| υ = kcat[ES] Vmax = kcat[E]t |
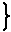 |
→ | υ/Vmax =[ES]/[E]t = r(飽和度) | (2) |
∴[ES] = r[E]t |
(3) | |||
| (3)式を(1)式に代入して, | ||||
| [S]=[S]t − r[E]t |
(4) | |||
(2)式からわかるように,υ/Vmax が基質Sによる酵素の飽和度rを表わしているので,酵素反応速度υを測定するということは,酵素Eへの基質Sの結合を速度を通してモニターしているということになる.ちなみに,ここでの考え方は酵素反応速度論におけるυ/[S]-υプロット (Eadie-Hofstee plot)やScatchard Plot (n=1)の場合と同じである.
Eadie-Hofstee の式
υ/[S] = Vmax/Km − υ/Km
Scatchard の式(n=1の場合)
r/[S] = 1/Km − r/Km
■酵素反応での拮抗阻害
●反応のスキームやモデルを書き下すということ
拮抗阻害のモデルは text には次のように書かれている.
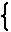 |
E + S 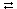 ES E + P ES E + PE + I EI |
・E,S,Pともにその存在状態はそれぞれ1種類しかない.E’などはない.
・EとSは1対1で結合し,ES中間体は1種類しかない.
・反応 E + S
ES,反応 ESE + P は可逆反応である.・Eは反応後,再生する.すなわちEは触媒である.
・反応を左→右,右→左と見ても同じである.SとPを入れかえても同じで,対称的である.
・SがEによらずPに変化することはできない.ESという中間体を経なければならない.S→Pという自発的反応はない.
・EとIは1対1で結合し,EI中間体は1種類しかない.
・反応 E + I
EI は可逆反応である.ここで,たとえば,阻害剤Iが基質Sの結合部位以外に結合し,その結果,酵素の構造が変化し,EIはSを結合できなくなってもよい.この場合もその実験結果は上の式で表される.「拮抗」というイメージと上の式で表されたモデルとは別である.上の式は「拮抗阻害」と我々が名づけた酵素の阻害現象もその1つとして含むだけである.繰り返すが,拮抗阻害ならば上の式で表されるが,逆に上の式は拮抗阻害のみを表しているわけではない.
したがって,L-B plot(実験結果)が図のようになったからといって,直ちに化合物Iは基質の結合部位に基質と競合して結合するとは結論できない.図の結果は確定証明ではないことに注意.EI複合体のX線結晶解析で,活性部位の基質結合部位に化合物Iが1分子結合し,酵素の他の部位には結合していないというデータがあれば,Iは拮抗阻害剤であると結論されよう.こういう意味において,kineticsには限界があることを明確に意識しておくことは重要である.他の実験方法においても同様である.それぞれの方法の限界を知るべし.
●基質の結合部位に結合するのに「拮抗阻害」を示さない例
1つの基質Sが1つの生成物Pに変化するという酵素反応(S → P)は非常に少ない.あるいはないといっても過言でない.多くの酵素反応では基質,生成物ともに2つ以上である.
さて,ここで2基質反応系の1つであるlactate dehydrogenase (LDH)を考える.
ピルビン酸 + NADH + Hイオン(H+)
乳酸 + NADイオン(NAD+)ピルビン酸の還元による乳酸の生成反応を見ている場合(ピルビン酸 → 乳酸),オキサミン酸oxamateはピルビン酸と「拮抗」する阻害剤である.しかし,同じピルビン酸の結合部位に結合するシュウ酸oxalateは「拮抗阻害」をせず,いわゆる「非拮抗阻害」をする.
LDHの反応機構は定序逐次機構(ordered sequential mechanism)であり,ピルビン酸とoxamateはE-NADHという同じ酵素形に結合しうるが,oxalateは異なる酵素形であるE-NAD+のピルビン酸結合部位に結合する.oxalateは非拮抗阻害を示すが,その結合部位は基質であるピルビン酸と同じ場所である.
逆に,乳酸の酸化(乳酸→ピルビン酸)の反応を観察しているときには,oxalateは乳酸と拮抗する.
「拮抗」という言葉は,基質と阻害剤が酵素の同じ部位を取り合うというイメージに対して,人がその現象に勝手につけたものであることに注意しよう.
■反応系や応答のモデルをたてたとき,それが意味すること
実験的研究においては先に式があるのではなく,まず第一に実験データ・実験事実があるのである.これは大切なことである.実験事実を積み重ねて,モデル・スキーム・式を作り上げる.そのモデル・式から今度は演繹的に予想・予測を導き出す.それをさらに実験的に確かめる.こうして,いろんな実験事実から最終的にモデルを書き下す.その時点でその研究は90%以上終わったも同然である.
このことは何もkineticsに限ったことではない.たとえば,cell biologyのような分野で定性的な実験をしたとする.ある化合物Xの影響を培養細胞で観察し,ある細胞応答の増大があったとする.その時間経過がlagを示しS字状に増加したのか,あるいはそうでなかったのか.このような実験から定性的にモデル(機構に関する考え=推論)をたてる.それに基づいて,定量的に考えを進め,実験・研究を展開する.当然,モデルによってそのやり方は変わってくる.こうして,帰納・演繹を繰り返しながら,スキーム・モデルを精密にしていき,研究を展開し,自然を解明していく.
●「 E + S ES → E + P 」というモデルの意味すること
ES E + P 」というモデルの意味すること|
|
先頭へ |
Last Updated 2005/8/18